Flavio Mattavelli.
Marginella sebastiani
nelle sue 5 varietà principali.
Oltre alla varietà tipica (1979),
distinzione delle varietà M. sebastiani millepunctata, tenuipunctata e “lellia” Mattavelli, 2022.
Quest’ultima in 2 subvarietà, lellia tondapex e
lellia acutapex.
Le “specie” per brevità potrebbero essere
talora scritte con singole lettere di una mia
simbologia sintetica, come elenco indicato alla fine del Post scriptum (es. “sebastiani” = S).
Quando
scrivo un nome specifico virgolettato
intendo trattare perlopiù di una vera specie biologica,
ma anche talora di una “supponibile specie”, che potrebbe essere in alternativa
un ipotetico “klepton”,
o anche soltanto una “varietà” di una vera specie o di un klepton.
“Varietà” è intesa sinonimo di morfo
di una popolazione, o anche solo di una forma individuale di un esemplare.
Non
intendo trattare di “varietà” subspecifiche o subkleptoniche
senza appello, cioè lascio la strada aperta alle 4 possibilità biologiche di classificazione di un esemplare ignoto:
vera specie/vero klepton/varietà o subvarietà di sp. o di kl. (già
noti o nuovi)/ibrido sterile.
Descrizione originale di M. sebastiani.
Spesso “sebastiani” in diversi riferimenti esteri si trova
scritto come “sebastiana”,
ma si tratta di refusi. M. sebastiani Marche-Marchad,
I. & Rosso, J.-C., 1979 fu descritta per la prima volta sul
Bollettino Malacologico UMI 1979, nell’articolo intitolato:
Une nouvelle marginelle
de la côte occidentale d'Afrique: Marginella sebastiani sp.
nouvelle (Gastropoda, Marginellidae). Bollettino Malacologico 15 (7-8); pag.
197-208.
All’interno delle popolazioni della specie M. sebastiani esistono almeno
5 notevoli varietà morfologiche specifiche, in generale tutte
chiamate “sebastiani”,
sulle quali sarebbe meglio indagare anche geneticamente. Io mi limiterò ad un’analisi morfologica, in base alla quale la specie M. sebastiani può formare un interessante Complex,
ricollegabile direttamente con il magico M. glabella Complex, allargato in questo sito Web a
tutte le marginelle del West Africa simili a M. glabella Linneo, 1758/1767, che è la specie tipo del Genus Marginella Lamarck, 1799.
In Appendice potete vedere la composizione di
tutto un M. glabella & sebastiani
Complex massimamente allargato a tutte le marginelle a loro simili, con le relative sigle
sintetiche.
Ripasso.
La varietà descritta come tipica di M. sebastiani mi appare poco più slanciata rispetto alla
immediatamente successiva foto di “lellia”, tuttavia con simile struttura di base.
La struttura
conchigliare di M.
sebastiani sebastiani,
cioè tipica, è precisamente come l’olotipo (foto v. avanti). La spira apicale
di quasi tutte le S normalmente non appare ribassata, ma nemmeno elongata
rispetto all’olotipo, inoltre l’ultimo giro è ben allungato, rispetto alla
lunghezza totale.
Le tipologie di denticolature interne del margine possono variare da
assenti a forti, con margine da stretto a ingrossato, come pure i tipi di puntinature della livrea possono
variare notevolmente, vedere avanti apposita trattazione dettagliata. Tuttavia
tra le S in media le “lellia”
adulte sono più dentate delle altre S e di P, inoltre le “lellia” in generale hanno conchiglie più
corpose e con puntinature più grandi e meno fitte (cioè a macchie rade).
L’analisi delle puntinature condurrà anche alla distinzione di 3 varietà base
di M. sebastiani
quasi tipiche, oltre a 2 subvarietà più
appariscenti della forma “lellia”.
Le
flammule assiali subsuturali chiare sono in generale assenti in M. sebastiani, tranne che in esemplari gerontici, ove non sono mai strette e lunghe. Ricordo che la principale differenza
rispetto alla simile M. pseudosebastiani Mattavelli, 2001 (= P, foto v. avanti) è appunto l’entità e
la forma delle flammule subsuturali, molto sottili e
lunghe solo in P, ove sono sempre
presenti.
Prime
evidenze di “lellia”,
però foto in esemplari
giovanili.


Nel
2006, nel mio Atlante
del Complex in questione, già denominai “lellia” una allora ipotizzata varietà di M. sebastiani
Marche-Marchad & Rosso 1979, di cui riporto le
foto di 2 esemplari giovanili, lunghi 41 mm.
Di “lellia” avevo
già scritto anche nel forum Malachia WMMCat 2007 e
nell’articolo Universo lumperia.
I punti,
ma nel caso di “lellia”
meglio dire macchie, evidenti sulla livrea di tali conchiglie sono da confrontare,
oltre che con quelli di M. pseudosebastiani, anche con quelli di diverse altre marginelle
più o meno definibili “ghepardiche”.
Derivatio nominis di “lellia”.
Con il senno
di poi, forse sarebbe stato meno confondibile introdurre, anzicché
il nome “lellia”,
la denominazione “pseudogoodalli”
(in confronto alla vera M. goodalli, v.
avanti) , come sinonimo (invalido)
della dubbia varietà di S in questione, in considerazione del fatto che ho
introdotto nel 2018 la diversa denominazione di “lellae”
per un’altra dubbia varietà/specie/klepton del
medesimo Complex allargato a tutte le “specie” di marginelle del West Africa tra loro simili (vedere www.pseudospecie.it/tris.htm
). Forse l’errore fu nel 2018, ma del senno di poi sono piene le fosse. Infatti
la varietà/specie M. (kl.?) pseudoglabella
lellae Mattavelli, 2018 ha ricevuto una
denominazione pure derivata da Lella, ma ovviamente ho mutato le 2 desinenze
latine, nell’ipotesi che potrebbe trattarsi di 2 effettive specie/klepton comunque diverse tra loro. Ancor oggi opto per la
confondibile coesistenza dei nomi “lellia” e “lellae”, in considerazione dell’osservazione che entrambi
tali nomi sono derivati dal soprannome Lella di mia moglie Graziella e mai
cambiare i nomi dedicati alle mogli.
Prologo, distinzione da M. goodalli e separazione di M. pseudosebastiani.
La prima
cosa che si nota osservando le conchiglie del Marginella glabella & sebastiani
Complex sono i punti chiari, talora quasi macchie, da
2 a 4 mm di diametro, quando ben distinti su sfondo ambrato o vinato. E’ una
livrea puntinata di solito a distribuzione casuale, con finezze e fittezze dei
punti diverse, però non caratteristiche di una sola specie biologica. Tant’è
che M. sebastiani
prima del 1979 fu confusa con M. goodalli, pure similmente punteggiata.
In
realtà M. sebastiani
può confondersi con M. goodalli soltanto nella varietà M. sebastiani lellia,
mentre nelle altre varietà S è più facilmente distinguibile da O.
 Ho
ripreso da Pinterest (foto a sinistra) una M. goodalli G. B. Sowerby,
1825, pressoché tipica, dimensioni visivamente comparabili nel confronto con le
foto di M. sebastiani
lellia.
Ho
ripreso da Pinterest (foto a sinistra) una M. goodalli G. B. Sowerby,
1825, pressoché tipica, dimensioni visivamente comparabili nel confronto con le
foto di M. sebastiani
lellia.
Occorre
considerare la reale separazione di M. sebastiani da M. goodalli, soprattutto in base alle diverse strutture
scheletriche, corroborate perlopiù dalle diverse taglie conchigliari.
M. goodalli è più
piccola, più tozza, più dentata, più larga, più conicheggiante
e spallata nell’ultimo giro, con l’apice più ribassato e stretto sulla spalla,
in paragone alle S “lellia”,
ed a maggior ragione in paragone alle S quasi tipiche etc., che sono più
affusolate.
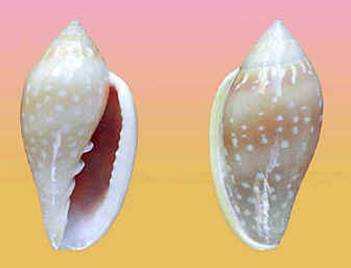 La mia
prima marginella “simil-sebastiani”
fu quella individuata come M. pseudosebastiani Mattavelli, 2001 (foto a sinistra), ora riconosciuta da WoRMS
(World Register of Marine Species)
come specie indipendente, sebbene ancor’oggi
morfologicamente confusa da qualcuno con
S, per non parlare in generale di un’apparente indistinguibilità genetica da M. glabella atlantidis,
come ritenuto da BOLD
(Barcode of Life Data Systems).
La mia
prima marginella “simil-sebastiani”
fu quella individuata come M. pseudosebastiani Mattavelli, 2001 (foto a sinistra), ora riconosciuta da WoRMS
(World Register of Marine Species)
come specie indipendente, sebbene ancor’oggi
morfologicamente confusa da qualcuno con
S, per non parlare in generale di un’apparente indistinguibilità genetica da M. glabella atlantidis,
come ritenuto da BOLD
(Barcode of Life Data Systems).
Analisi morfologica di “sebastiani”.
Riguardo
alle popolazioni della specie M. sebastiani, con la maggior conoscenza di svariati
esemplari, oggi per me, pur prudenzialmente “lumper”,
occorre riconoscere almeno 2 popolazioni
estreme di M. sebastiani,
con caratteristiche morfologiche diverse e probabilmente con biologia
indipendente, a parte poi i casi di esemplari morfologicamente intermedi tra M. sebastiani
e le specie più prossime, cioè i cosidetti ibridi, o forse popolazioni klepton ibridogeniche.
Distinguerò
dunque M. sebastiani Marche-Marchad & Rosso, 1979 “tipica”, altrimenti denominabile M.
sebastiani sebastiani,
e M.
sebastiani lellia
Mattavelli, 2022, intendendo la seconda “varietà” non già come una vera nuova
“specie/klepton” indipendente, ma come una “varietà”
morfologica diversa e diversificata prudenzialmente solo come nuova varietà,
non ancora riconoscibile biologicamente come una vera nuova specie
indipendente, oppure in alternativa un nuovo klepton
indipendente.
Si vedrà
che “lellia”
a sua volta è suddividibile in 2 subvarietà
distinguibili, ma unificabili in essa.
 Preciso
che è ambiguo affermare storicamente quale sia proprio la forma tipica di M. sebastiani
Marche-Marchad & Rosso, 1979, sebbene l’analisi
del documento originale (Bollettino Malacologico UMI 1979) mi induca a ritenere
tipica quella di scheletro più allungato
ed in genere di taglia maggiore, con sul dorso puntini leggermente più
piccoli e con il labbro esterno del margine meno evidente, in ciò corroborato
dalla foto a sinistra dell’olotipo di M. sebastiani,
lungo 55,4 mm, presente al MNHN di Parigi (foto Caballer).
Nello stesso Bollettino UMI 1979 la distinzione tra i 2 morfi di M. sebastiani,
tipico e “lellia”,
non è comunque evidenziata.
Preciso
che è ambiguo affermare storicamente quale sia proprio la forma tipica di M. sebastiani
Marche-Marchad & Rosso, 1979, sebbene l’analisi
del documento originale (Bollettino Malacologico UMI 1979) mi induca a ritenere
tipica quella di scheletro più allungato
ed in genere di taglia maggiore, con sul dorso puntini leggermente più
piccoli e con il labbro esterno del margine meno evidente, in ciò corroborato
dalla foto a sinistra dell’olotipo di M. sebastiani,
lungo 55,4 mm, presente al MNHN di Parigi (foto Caballer).
Nello stesso Bollettino UMI 1979 la distinzione tra i 2 morfi di M. sebastiani,
tipico e “lellia”,
non è comunque evidenziata.
Distinzione e definizione di “lellia” Mattavelli,
2022, in 2 subvarietà.
 Definisco
“lellia” sintipica ognuno
dei 2 esemplari esposti a lato, entrambi lunghezza 40 mm, senza dare
particolare importanza alle puntinature a macchie delle loro livree conchigliari, salvo notare che hanno sempre relativamente
poche grandi macchie chiare ad interassi tra i centri delle macchie perlopiù
elevati oltre il doppio dei diametri delle macchie.
Definisco
“lellia” sintipica ognuno
dei 2 esemplari esposti a lato, entrambi lunghezza 40 mm, senza dare
particolare importanza alle puntinature a macchie delle loro livree conchigliari, salvo notare che hanno sempre relativamente
poche grandi macchie chiare ad interassi tra i centri delle macchie perlopiù
elevati oltre il doppio dei diametri delle macchie.
Non
darei importanza nemmeno alle diverse denticolature
interne dei margini (denticolature comunque più
evidenti di quella in apparenza mancante nell’olotipo di S fotografato da Caballer), mentre notare piuttosto che la struttura scheletrica dei 2 sintipi di “lellia” è diversa nella spira apicale e nella forma
boccale, con ben diverse spallature dell’ultimo giro, subito sotto la sutura
della cuspide.
Pertanto
attribuisco all’esemplare a sinistra il nome di “lellia tondapex”, mentre all’esemplare a destra il nome di
“lellia acutapex”,
intendendo la presenza di 2 sub varietà limite della stessa F. Naturalmente
esistono altri esemplari intermedi di F, che non sono né “tondapex”
né “acutapex” (v. foto in Conclusione). La coloritura
do fondo degli esemplari freschi può essere anche più rosso vinata di quelli
fotografati.
Alcune altre conchiglie quasi “lellia”.

 Riportai nella Guida
anche le foto di 2 esemplari "stocky &
dark" sebastiani, 45 + 38
mm, con margine dentato (non molto), qui ricopiate. Queste conchiglie "tozze e scure" sono meno
affusolate, più robuste e corpose della M.
sebastiani sebastiani
tipica, che appare invece con il margine più sottile. Per le dimensioni e le caratteristiche direi che si tratta di morfi quasi “lellia acutapex”, tuttavia con
macchie chiare più piccole e ravvicinate.
Riportai nella Guida
anche le foto di 2 esemplari "stocky &
dark" sebastiani, 45 + 38
mm, con margine dentato (non molto), qui ricopiate. Queste conchiglie "tozze e scure" sono meno
affusolate, più robuste e corpose della M.
sebastiani sebastiani
tipica, che appare invece con il margine più sottile. Per le dimensioni e le caratteristiche direi che si tratta di morfi quasi “lellia acutapex”, tuttavia con
macchie chiare più piccole e ravvicinate.
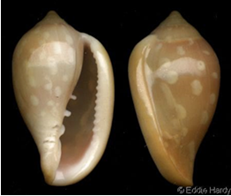
Dal sito
Web di Eddie Hardy, ora accessibile tramite il sito Conchology Inc., ho copiato qui quel che si potrebbe chiamare ancora “lellia” quasi
tipica, o meglio “lellia tondapex”,
però troppo inscurita nel colorito di fondo della livrea, comunque abbastanza ben
denticolata sul margine ingrossato (esemplare adulto) e presentante la spira
apicale ribassata.
Le
differenze morfologiche rispetto all’olotipo di “sebastiani” sono abbastanza evidenti.
Ciò che
più distingue tale “lellia
tondapex” è lo scheletro più tondeggiante nella
zona apicale. L’allungamento totale conchigliare è poco minore di quello dell’olotipo di M. sebastiani e di tutte le S dei prossimi
capitoli, cioè mediamente 1,80 < 1,96 circa.
L’allungamento
boccale è invece maggiore di quello
delle S quasi tipiche e delle “lella
acutapex”, che lo ha simile a quello delle S quasi tipiche.
Approfondimento sulle forme di M. sebastiani
non riconducubili alle precedenti “lellia”.
In
questa categoria rientrano innanzitutto l’olotipo di S e le S che verranno
chiamate quasi tipiche.
Nella
mia Guida o Atlante Internet
del 2006 avevo anche messo le foto recto e verso di una forma di "mixed
& elongate" sebastiani, qui di seguito
ricopiate, intendendole di un morfo ben ricollegabile al tipico S.
Esistono differenti varietà allungate di M.
sebastiani con caratteristiche tra loro miscelate
quasi tipiche, ma talora intermedie con le specie vicine e con “lellia”.
Ecco
dunque un esemplare quasi tipico di M. sebastiani,
Guinea, 52 mm, che potrebbe anche confondersi con M. pseudosebastiani.
Invece M. pseudosebastiani presenta
solitamente i puntini più piccoli e leggermente allineati, in uno sfondo più
chiaro. La taglia di M. sebastiani è
solitamente più piccola di M. pseudosebastiani, che inoltre presenta sempre le flammule
assiali subsuturali distintive, che nelle
foto seguenti mancano.
La denticolatura del margine dell’esemplare 52 mm è insolitamente ben presente, però in altri
esemplari simili, compreso l’olotipo, ed in tutti i prossimi fotografati, è in
generale assente.


Confronto delle caratteristiche
morfologiche.
La denticolatura
del margine interno delle “lellia”, pur con notevoli casi ambigui, perlopiù giovanili
sdentati, nella maggioranza degli
esemplari adulti appare in generale più evidente in “lellia”,
piuttosto che nelle tipiche e quasi tipiche M.
sebastiani sebastiani
adulte.
Nel
confronto delle puntinature di S e “lellia” fondamentalmente
si possono distinguere i seguenti 4 tipi di punti chiari. Poi esistono forse
dei casi di ibridazione SXP o SXG o SXR o SXL, da studiare a parte.
- Puntinatura a finezza e fittezza ghepardica
grossolana, cioè più fitta e più fine dell’esemplare 52 mm,
casi concretizzabili nella varietà da me chiamata M. sebastiani millepunctata (ma in realtà
questa tipologia è soltanto ben multi puntata), casi abbastanza rari.
- Puntinatura più rada a punti di pari finezza del caso precedente,
o con macchiette di diametro un poco maggiore (cioè come il morfo 52 mm):
non darei un nome definitivo a queste comuni varietà “mixed & elongate” quasi tipiche di S, considerandole casi intermedi tra l’essere o ben multi punctati oppure
pauci punctati, o
meglio pauci e insieme parvi punctati,
con punti di diametro tra 4 e 2 mm.
- Puntinatura quasi assente, punti generalmente di diametro
medio-piccolo, casi proprio a punti parvi e insieme pauci
puntati, che verranno denominati M. sebastiani
tenuipunctata.
- Puntinatura molto rada, con punti degenerati in grandi macchie.
E’ una prerogativa sia di “lellia”
che tendenzialmente degli esemplari quasi
tipici di M. sebastiani, sebbene questi ultimi
rientrino talora, come puntinature, nei casi parvipunctati
o paucipunctati, mentre “lellia”
non presenta quasi mai punti piccoli, e ha macchie perlopiù abbastanza
grandi e rade, quasi sempre leggermente più grandi e meno numerose di quelle
delle forme quasi tipiche e della tipica M. sebastiani sebastiani.
Le denticolature e le puntinature vanno viste associata alla struttura scheletrica, che è o con profilo più o meno come “lellia”, cioè
robusta ad apice perlopiù solitamente
non ribassato (casi “acutapex”), ma talora ribassato nella sub varietà
“tondapex”,
oppure la struttura è come nei casi quasi tipici di S, etc., cioè la struttura
è più slanciata, ad apice alto, di
solito non eccessivamente elongato. Le “lellia” sono di taglia generalmente minore. Diciamo che
solo “lellia tondapex”
risulta con spira apicale ribassata, mentre le S perlopiù hanno la spira
apicale più rialzata, ma non elongatissima.
Riguardo
alle coloriture degli sfondi delle
livree conchigliari notare che gli esemplari più
freschi di “lellia”
tendono di sfondo livrea al rosso vinato, mentre quelli più “stagionati” in
musei o in collezioni private tendono al color salmone ingiallito o brunito; ad
esempio vedere esemplari come già rappresentati nell’Atlante 2006
e Universo lumperia, laddove le forme quasi tipiche di M. sebastiani
in generale possono raggiungere coloriture mattone più brunito delle “lellia”, che
sono invece color mattone piuttosto tendente al rosso, talora “pompeiano”.
Quindi le
varietà limite non ibridate, cioè i casi estremi con caratteristiche specifiche
come sopra elencate, sarebbero ascrivibili in totale a 5 tipologie non tutte accumunabili, es. tra loro “lellia” non
mi pare che presenti le puntinature ghepardiche della
“S millepunctata”,
tranne che in alcuni casi “dark &
stocky” = quasi “lellia acutapex”.
Riassumendo, in totale riconosco 2 + 3 = 5
“varietà” principali di M. sebastiani:
- “lellia acutapex”
(come nelle foto viste in precedenza)
- “lellia
tondapex” (come nelle foto viste in
precedenza)
- S
tipica e quasi
tipica (perlopiù puntata come olotipo, ma anche come il precedente esemplare
52 mm e come i primi 2 esemplari quasi tipici seguenti)
- S
millepunctata
(come descritta nel link e v. avanti casi anomali)
- S tenuipunctata
(cioè S sia parvi che pauci punctata
insieme, v. avanti casi anomali).


A
sinistra S quasi tipica tendente pauci puntata a macchie.
A destra
S quasi tipica tendente multipunctata a punti più parvi, ma non ancora definibile S millepunctata
“ghepardica”, in quanto si notano molti
spazi vuoti tra le macchiette del dorso conchigliare
(foto prese dalla pagina
di Hardy nel sito Conchology Inc., foto rispettivamente ©dei Sig.ri Poppe e ©di Lowtide Shells).
Casi anomali multipunctati
ed all’opposto casi di M. sebastiani tenuipunctata.
Faccio
ancora seguito all’articolo Un
salto nei retini conchigliari “ghepardici”
La
varietà M. sebastiani
millepunctata, cioè retinata stocastica quasi
uniformemente “ghepardica”, è abbastanza rara.
Tranne “lellia”, quasi
tutte le S sono confondibili con M. pseudosebastiani, però “S millepunctata” è più confondibile per via dei punti
fini e molto fitti, ma non è confondibile per il colore di fondo delle
conchiglie. S è generalmente assai più scura, talora rosseggiante al marrone,
piuttosto di P, che invece tende maggiormente al giallino-rosato luminoso.
Inoltre,
come già scritto, “lellia”
tende maggiormente al rosso “pompeiano”, tranne i casi “dark”.
Questa regola dei colori ha naturalmente le
dovute eccezioni, vedere ad es. l’olotipo di S giallino per natura, per di più
anche quando i colori potrebbero spegnersi nel tempo, per una lunga
conservazione museale.
Inoltre
esistono pure esemplari “rusty”, color ruggine, rimando al qui
presente link.
Infine
notare che casi di S paucipunctati tendono di sfondo al
color sabbia gialla, vedere successivamente l’immagine della forma S tenuipunctata.
Ecco
dunque prima subito una varietà di S ben colorita mattone (foto © Alboran Shells in Hardy), con punti chiari non definibili piccoli, ma,
essendo molto numerosi e abbastanza fitti, definibili “ghepardici”,
pertanto tale varietà multi punctata potrebbe essere una
forma “S millepunctata”, anche se
le macchiette non sono numerosissime ed uniformi, come nell’esemplare da cui è
nato il nome di tale varietà. Si nota
anche un certo allineamento delle macchiette, tendente alla livrea di P, tranne
il color mattone del fondo. Penso che l’esemplare sia S, pur se la discutibile
presenza di piccole flammule subsuturali strette
farebbe tendere ad una M. pseudosebastiani a puntini grandi, oppure quanto meno
ad un ibrido SXP; oppure indurrebbe anche ad un ibrido SXR?
Si nota
anche un certo allineamento delle macchiette, tendente alla livrea di P, tranne
il color mattone del fondo. Penso che l’esemplare sia S, pur se la discutibile
presenza di piccole flammule subsuturali strette
farebbe tendere ad una M. pseudosebastiani a puntini grandi, oppure quanto meno
ad un ibrido SXP; oppure indurrebbe anche ad un ibrido SXR?
L’ibridazione
SXR è improbabile in quanto, pur non essendo da S la struttura del margine conchigliare, che è schiacciata dritta a centro margine,
come in alcune M. pseudodesjardini Le Béon,
2012 (= R), R ha una conchiglia comunque più allungata!
 Esistono
esemplari di S che all’opposto presentano pochissimi punti chiari ed anche
perlopiù piccoli (foto © Conchology Inc. in Hardy), su sfondo color sabbia gialla.
Esistono
esemplari di S che all’opposto presentano pochissimi punti chiari ed anche
perlopiù piccoli (foto © Conchology Inc. in Hardy), su sfondo color sabbia gialla.
Le
macchie, anzi i puntini chiari, sono qui quasi assenti e comunque più piccoli e
scarsi del solito, da introdurre una varietà
S parvi punctata & pauci
punctata insieme.
Per
abbreviare chiamerò questo caso ipopunctato M. sebastiani tenuipunctata.
Albinismo. Vedere
Fig. 16 e 17 dell’articolo specifico.
Senza
sbiancare completamente anche il fondo livrea, alcune ambiguamente credibili S
hanno macchie indefinite e spalmate su fondo livrea a fasce alterne
chiaro/beige (es. Fig. 17, con tendenza alle S quasi tipiche), oppure hanno
punti quasi assenti su fondo color cammello chiaro (es. “galindoi” Fig. 16, invero non “albineggiante”). L’esemplare isolato “galindoi” si può forse collegare
anche alla forma “problematica”,
intermedia tra G e V, quindi ora forse pure collegabile ad S, ma una rondine
non fa primavera.
Conclusione.
Soltanto
“lellia” (=
F, vedere Appendice) potrebbe aspirare ad un ruolo tassonomico superiore alla
varietà specifica, cioè essere candidata a possibile nuova specie/klepton, o quantomeno
potrebbe essere una sottospecie ambientale di S, se non essere un klepton, forse SXO, trascurando per rarità i casi
sopraindicati delle presunte S “albineggianti” e
scartando, per illogicità delle morfologie, possibili ibridi SXG, oppure SXV,
ove le specie ibridanti siano supposte quali parentali di “lellia” ipotizzata klepton. E’
un’ipotesi comunque troppo remota.
Per
assurdo, se F fosse specie/kl. separata, potrebbero
nascere ibridi interspecifici tra S e F, dei quali avremmo già caratteristiche morfologiche di ibridazione
in atto, esempio la denticolatura e la grossezza
del margine di qualche S quasi tipica, o talora “tozzezze”
di struttura intermedie tra quelle tipiche di S e F, con spire apicali perlopiù
di elevazione comune “normali”, cioè né ribassate come in alcune “lellia tondapex”, né rialzata in forme acute, intendendo
cuspidi “normali” come sono perlopiù nelle quasi tipiche S (es. come nel morfo "mixed & elongate" 52 mm),
per non parlare ancora di tipologie di puntinature intermedie assai comuni tra
S & F.
Le
diversità morfologiche esistono, ma non mi appaiono nettamente determinanti per
una separazione biologica specifica, dato l’elevato numero di individui con
caratteristiche comuni a varietà principali diverse. Pertanto non ritengo F una specie diversa da S.
Ogni singolo
esemplare individuale può presentare qualche caratteristica anomala della “subvarietà” di appartenenza, ogni “subvarietà”
può presentare caratteristiche comuni di individuazione morfologica con altre “subvarietà”, o addirittura con altre specie/kl.,
ovviamente senza pretesa di assoluta individuazione specifica, nel senso di
individuare esattamente la categoria biologica di appartenenza dell’esemplare
in esame.
Notare
che tutte le “lellia”
non superano 50 mm di lunghezza, mentre le forme “quasi tipiche di S” possono raggiungere 70 mm; ciò potrebbe far
pensare che le “lellia”
siano, un po’ tozze, forme nane e le
altre, poco più slanciate, siano forme
giganti della medesima S, magari entrambe sottospecie geografiche locali,
ma non ho informazioni decisive su tutte le provenienze, spesso sovente
indicate genericamente Senegal.
In
conclusione le taglie delle conchiglie sono un buon indizio, ma non una prova
della loro identità.
Anche la
coloritura di fondo non è sempre significativa, perché esistono “lellia”
aranciate, come alcune S quasi tipiche, anziché come le in precedenza descritte
“lellia”
vinate “pompeiane”.


Nelle 2
aranciate immagini soprastanti: a sinistra una “lellia”
41,7 mm, intermedia tra “tondapex
“ e “acutapex”; a destra una S quasi tipica 59,7 mm; provenienza
di entrambe generalizzata Senegal.
Gli
allungamenti totali sono circa 1,91 e 1,96, valore normale per le S tipiche, ma
assai elevato per la maggioranza delle “lellia”. L’allineamento di alcuni puntini della “lellia” 41,7 è pure anomalo.
 Altro
esemplare, ma proveniente dalla Guinea (foto Alboran Shells
rilevata da un'asta Web) lunghezza 44 mm, esempio di connubio morfologico tra
le varietà tipica della specie Marginella sebastiani e "lellia", perché ritengo tali
2 varietà subspecifiche e non 2 specie biologiche diverse, pur essendo talora distinguibili
in forme morfologiche separabili.
Altro
esemplare, ma proveniente dalla Guinea (foto Alboran Shells
rilevata da un'asta Web) lunghezza 44 mm, esempio di connubio morfologico tra
le varietà tipica della specie Marginella sebastiani e "lellia", perché ritengo tali
2 varietà subspecifiche e non 2 specie biologiche diverse, pur essendo talora distinguibili
in forme morfologiche separabili.
Quest'esemplare,
in prevalenza definibile “lellia”, allungamento 1,78, è notevole per lo
spessore del margine come le “lellia” adulte e
puntinatura quasi tipica di “lellia”, ma unitamente
ad una struttura conchigliare poco più esile, come le
forme quasi tipiche di S e di loro quasi pari coloritura intermedia.
Relazioni con altre specie del Complex super allargato.
Riepilogando
le tipologie di puntinature, mentre
i puntini chiari di M. glabella atalanta, G bordeaux, M. irrorata pseudoirrorata, pseudoglabella lellae, visayae pseudovisayae e pseudosebastiani
sono generalmente “ghepardici” sotto i 2 mm di
diametro (e ravvicinati), i punti delle M.
sebastiani definite quasi tipiche e soprattutto
delle M. sebastiani
lellia sono generalmente sopra i 3 mm di diametro
e distanziati tra loro anche ben oltre il diametro, determinando interassi
anche molto oltre 2 diametri.
Fino al
2001 le specie non fossili e note del M. glabella & sebastiani Complex, inteso super allargato a M. irrorata, goodalli, aurantia
ed alle altre marginelle del West Africa più simili a
M. sebastiani,
cioè allora solo M. desjardini,
erano allora descritte ufficialmente soltanto 6 (A, D, G, O, I, S).
Nel 2001
io buttai un sasso nello stagno introducendo P. Oggi quelle descritte in
cartaceo sono diventate 13, ma il loro numero è destinato a crescere, credo di
almeno 9 unità, comprendendo sia le vere specie, che i probabili klepton, e le loro varietà, cioè tutti quei morfi già oggi
distinguibili morfologicamente come morfi fra loro diversi, ma biologicamente
di assegnazione ambigua, cioè se riconoscibili come vere nuove sp./kl., oppure notevoli varietà, ma solo varietà subspecifiche.
Una di queste ambigue nuove “specie”
potrebbe essere Marginella (kl.?) lellia, ma credo sia solo
una bella “varietà” di S, leggermente meno allungata e di struttura
poco più robusta delle altre varietà tipica e quasi tipiche di S.
Flavio Mattavelli Release
14/09/2022 matta.a@tiscali.it

Post scriptum Maggio 2023
Morfi
inclassificabili
Attorno alle 5 principali varietà di S
gravitano alcuni singoli esemplari che mi è difficile definire varietà di
specie, ma mi è altrettanto difficile definire ibridi biologici (in realtà sono ambigui “ibridi” solo morfologici), cioè
potrebbero essere, oltre che varietà di specie, forse ibridi veri o addirittura
ibridi klepton (ibridogenici)
di specie già note, escludendo la possibilità che potrebbe trattarsi di specie
nuove, per via della scarsità pratica degli esemplari rinvenuti (il che in
teoria non importerebbe).
Nel seguito tutte le scritture riguardanti
gli “ibridi” sono da intendersi valide anche se scritte al contrario, es. SXP =
PXS.
Ho già supposto un caso che potrebbe essere
SXG, ma anche SXV o VXG (?) nell’Appendice all’articolo Un salto nei retini ghepardici.
Ho trovato altri casi di esemplari che
riguardano perlopiù solo i morfi PXS (vedere avanti al punto 1). Altri casi
ancora di diversi morfi punteggiati verranno trattati al punto 2, imputandoli a
supposti ibridi SXL, o meglio SXE *.
Si può dunque ricollegare tali esemplari in un
gruppo SGPL, o meglio SGPE *, più magari V.
In tale gruppo possono esistere anche
ibridi non a base parentale S, ma somiglianti ad S, esempio LXG (o meglio EXG
*), oppure PXG o PXL (o meglio PXE *), nonché ibridi eventualmente riferibili
anche a V. Gli ibridi riferibili a V determinerebbero un allargamento del
gruppo da studiare, tuttavia possono rientrare forse in varietà di V in parte
già trattate nell’articolo definitivo di V,
quindi vengono momentaneamente trascurati, ma ci sarà un cenno al punto 3 di
questo Post scrptum.
Inoltre, in ultima analisi per le puntinature ghepardiche, alcuni morfi sono forse anche collegabili a Marginella pseudodesjardini
= R, con puntinature ben più grandi dei precedenti, da far pensare a
possibili SXR. Se si volesse allargare il gruppo SGPL si dovrebbe quindi
aggiungere anche R.
A fronte di 5 specie parentali, potrebbero
esserci 10 ibridi, derivati dalle combinazioni logiche dei loro collegamenti,
ad esempio potrebbero in teoria esistere:
SXP, SXL, SXG, PXL, PXG, LXG, RXS, RXP, RXL,
RXG.
Tuttavia non è detto che esistano
tutti questi ibridi teorici ed al momento trascuriamo
pure R ed eventuali suoi ibridi comunque intesi. Infatti non ho trovato tutti quegli
esemplari che potrebbero esserci come ibridi teorici, sulla base del numero di
specie considerate.
Volendo restringere il discorso ai morfi che appaiono in pratica più simili a
S, per cominciare fisserei l’attenzione solo sugli ibridi morfologici delle
4 specie:
1.
Marginella sebastiani = S
2.
Marginella pseudosebastiani = P
3.
Marginella pseudoglabella = L, ma
sarebbe meglio considerare E = L lellae *
4.
Marginella glabella = G, ma
sarebbe indispensabile considerare anche V = Marginella visayae, senza considerare altre
presunte “specie” o sotto specie, in particolare varietà di G, come ad esempio N = G atalanta, come introdotta alla
pagina Revisione di M. glabella,
colà Foto 12 B.
Per alcune foto delle forme tipiche di
base delle specie sopracitate riferirsi all’Atlante.
* Tra i
morfi punteggiati più riconducibili a LXG
faccio notare che M. lellae (= E, nel link Fig. 1 & 2) è stata
da me considerata forma di M. pseudoglabella,
come pure M. atalanta (= N, nel link Foto 12
B) è stata considerata forma di M. glabella.
Insisto sull’attribuzione non separata da L
& G di questi 2 morfi E & N, attribuzione che potrebbe essere
biologicamente errata, ma viene nella presente analisi mantenuta per
semplicità, a livello di 2 varietà di specie, allo scopo ridurre in prima
approssimazione il numero teorico dei possibili ibridi specifici.
L’analisi
può partire dalla considerazione che, a fronte delle suddette 4 specie GSPL, potrebbero esserci logicamente 6 ibridi
derivati dalle combinazioni teoriche
SXP, SXL, SXG, PXL, PXG, LXG.
Considerando anche V, con 5 specie base,
potrebbero esistere invece i 10 ibridi SXP, SXL, SXG, PXL, PXG, LXG, VXS, VXP,
VXL, VXG.
Però, a fronte di 6 specie base, le
combinazioni diverrebbero 15, mentre, a fronte di 7 specie base, le
combinazioni diverrebbero 21 e via conteggiando, secondo la formula n° ibridi finale = n° specie
base precedenti + n° ibridi della numerazione corrispondente al n° di specie
base precedenti.
Analisi di
alcuni morfi inclassificabili del gruppo GSPL collegabili alle caratteristiche di M. sebastiani (punti 1 & 2).
1. 

Esemplari a sinistra lunghezza 46 mm, a destra
lunghezza 48 mm, provenienza Mauritania, chiamati “pseudosebastiani” nel sito Alboranshells, ma con puntinature diverse da quelle
delle comuni P, cioè puntinature tendenti a quelle di S…
Direi che potrebbe trattarsi di 2 poco comuni ibridi
morfologici SXP, che potrebbero essere stati raccolti al confine tra
Mauritania e Senegal, zone ambientali più comuni rispettivamente per P e per S,
non negando quindi la possibilità di un’effettiva ibridazione sterile, ma che
potrebbe essere anche fertile (= klepton), qualora
tale ipotesi fosse suffragata dal ritrovamento di numerosi altri esemplari
simili, cosa che al momento non mi pare.
Notare che anche un’alternativa
identificazione come n-esima varietà di
P, oltre a quelle già fotografate in altro
articolo, non andrebbe scartata, mentre scarto l’idea di varietà di
S, perché sono presenti le flammule assiali subsuturali
caratteristiche di P, maggiormente nell’esemplare 46 mm!
In conclusione i 2 esemplari del punto 1 sono
abbastanza inclassificabili.
2. 
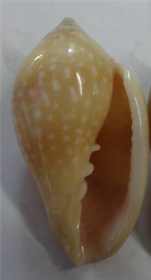
Ecco un altro esemplare della Mauritania, lunghezza 47,3 mm (foto del sito
Nautilus, colà dichiarato “pseudosebastiani”),
potrebbe essere proprio una n-esima
varietà di P o in alternativa un ibrido
PXS, ma, avendo una puntinatura tendente al ghepardico, ed
apparendomi quasi privo di flammule assiali subsuturali,
l’esemplare potrebbe essere anche una S varietà millepunctata.
Tuttavia quest’esemplare sfugge troppo verso L lellae = E, da
far pensare ad una possibile ibridazione
SXL, o meglio SXE (oppure anche, meno probabilmente per la mancanza di
flammule assiali subsuturali, PXE)*.
Tale ultima idea è rafforzata anche
dall’elevato allungamento dell’esemplare in questione, allungamento che
indurrebbe ad escludere qui, dall’eventuale ibridazione parentale base S,
riguardo all’altro genitore escludere la specie G e forse escludere anche V,
mentre abbiamo visto che in un altro esemplare (Zaurshells, nell’Appendice
all’articolo Un salto nei retini conchigliari ghepardici)
tale ibridazione può manifestarsi, nell’ambiguo ibrido SXG o SXV, se non GXV…
Analisi di
alcuni morfi inclassificabili del gruppo GSPL non collegabili alle caratteristiche di M. sebastiani (punto 3).
Ora occorrerebbe aggiungere la presenza
di M. visayae
e si apre un vaso di
Pandora riguardo alle punteggiature
ghepardiche di esemplari morfologicamente ibridi, analisi che in verità
riguarda maggiormente specie diverse da M.
sebastiani.
Già nell’Atlante era
apparso il problema di esemplari ambigui nel tetraedro di vertici G,S,P,L, vedere colà
la Fig. 16, ritenuta forse di una P giovanile, figura dove però veniva e
viene esclusa una possibile ibridazione di S con le altre specie del gruppo, le
quali invece possono ibridarsi tra loro e con V.
Vediamo un esempio di un morfo ambivalente a identificazione molto incerta, praticamente inclassificabile.
3. L’esemplare
seguente è lungo 37,3 mm, provenienza Dakar, classificato P dal Dealer
(Nautilus).
Per me l’identificazione è errata, in quanto
l’esemplare mi appare un ibrido morfologico non ben classificabile, potrebbe essere
tutto tranne P ed anche tranne S.
Avevo già trattato casi analoghi (vedere nel
link il capitolo Ibridi PXGXS?), ma allora non consideravo L, o meglio E*, e non
consideravo ancora V. Oggi L & V sono “specie” effettive, da non
trascurare, inoltre P & S in questo caso credo che non c’entrino affatto,
per i motivi seguenti.

 Se si esclude che l’esemplare 37,3 mm sia una
varietà di S o di P e se si esclude che possano esserci ibridazioni con S (per
via dei puntini troppo piccoli) o
con P (per via del relativamente troppo
basso allungamento conchigliare), potrebbe quindi
trattarsi di un ibrido nel triangolo GEV,
cioè potrebbe essere VXE o VXG (o
meglio VXN o meglio ancora YXN) o GXE oppure meglio NXE *.
Se si esclude che l’esemplare 37,3 mm sia una
varietà di S o di P e se si esclude che possano esserci ibridazioni con S (per
via dei puntini troppo piccoli) o
con P (per via del relativamente troppo
basso allungamento conchigliare), potrebbe quindi
trattarsi di un ibrido nel triangolo GEV,
cioè potrebbe essere VXE o VXG (o
meglio VXN o meglio ancora YXN) o GXE oppure meglio NXE *.
Nel gruppo GSPLV ho escluso S&P e ho
sostituito L con E, poi ho messo N al posto di G e Y al posto di V, perché le
puntinature di E, N, Y (considerate sottospecifiche di L, G, V) sono più vicine
alla punteggiatura dell’esemplare in questione.
Escluderei che possa trattarsi solo di una
varietà di V, cioè Y = M. visayae pseudovisayae, perché
i puntini sono troppo grossolani per una Y (invece per una vera Y vedere questo link).
Al contrario non escluderei che, per la
puntinatura quasi tipica di E, probabilmente potrebbe trattarsi solo di una E =
M. pseudoglabella
lellae nana, oppure in
alternativa di una N = M. glabella atalanta,
siccome N & E hanno puntinature quasi identiche, a parte le dimensioni
di E > N. Probabilmente staremmo
quindi osservando soltanto una varietà
di “specie” singola, ma ignota.
Dunque morfologicamente
non possiamo sapere di quale “specie” (ibrido o specie) si tratti!
Avendo già escluso S dall’analisi ibridatoria, tutto ciò non significa che non potrebbero
esistere ibridi LXP = EXP, GXP = NXP, VXP = YXP, ma si tratterebbe di una diversa indagine coinvolgente P, nel
supposto tetraedro GPLV, o meglio NPEY.*
Tuttavia ho già notato che la specie P ha
l’allungamento conchigliare maggiore di quello
dell’esemplare 37,3 mm, dunque per l’esemplare in questione il discorso ibrido
di P è decaduto.
* Osservo che sostituendo E a L, N a G e Y a V
si ottengono gli ibridi più probabili per l’esemplare in questione 37,3 mm, per
via delle puntinature più simili a quelle di E, N, Y, tuttavia logicamente
potrebbero esistere, senza coinvolgere P
& S, nel gruppo LEVYGN, qualora supposto costituito da 6 “specie”
distinte, ben 15 possibilità di ibridazione (combinazioni di specie), cioè:
GXV, GXL, LXV, NXY, NXE, EXY, GXY, GXE, YXL,
NXV, NXL, EXV, LXE, YXV, GXN.
Invece se si pone E=L, N=G, Y=V ovviamente ci
si riduce al triangolo LGV = ENY, con solo 3 reali combinazioni di ibridazione,
cioè: GXV = NXY, GXL = NXE, LXV = EXY.
In quest’ottica gli ultimi 3 ibridi
nell’elenco delle 15 combinazioni sono manifestamente solo virtuali, perché
tali ultime 3 combinazioni rappresenterebbero solo specie in varietà di morfi
solo specifici (LXE=L oppure varietà =E, YXV=V oppure varietà =Y, GXN=G oppure
varietà =N).
Riassunto
sulla mia simbologia sintetica.
Ecco l’elenco
alfabetico di tutte le “specie” recenti (non fossili) riconducibili al “glabella” Complex allargato al massimo grado di similitudine con G,
specie ricollegabili come galassie ruotanti attorno ad un nucleo centrale di
materia oscura (Complex in senso stretto costituito
solo dal trio “glabella/ visayae/
irrorata”), specie tutte del Genus Marginella Lamarck, 1799.
Le “specie” in questione recano affiancate lettere o sigle della mia simbologia
sintetica.
13 “specie” sono
attualmente già accettate da
WoRMS come vere
specie biologiche:
aurantia……………….……...…..A Lamarck, 1822
desjardini………………..……....D Marche-Marchad,
1957
glabella………………….…………G Linneo,
1758/1767
goodalli………………….…………O G. B. Sowerby,
1825
irrorata………………………..…….I Menke, 1828
lamarcki………………..………….K Boyer, 2004
(v. 5 confondibili marginelle
simil-glabella etc.)
mattavellii………………..……..M Cossignani,
2021 (v. Albinismo delle simil-glabella)
pseudodesjardini………….…..R
Le Béon, 2012 (v. Marginella pseudodesjardini)
pseudoglabella…………….……L
Mattavelli, 2018 (v. Descrizione comparativa)
pseudosebastiani……….……..P
Mattavelli, 2001 (MMM n°34, aprile 2001). Esistono almeno 2 varietà
principali di P, però non denominate singolarmente.
sebastiani ……….…………….….S Marche-Marchad
& Rosso, 1979, con almeno 2 varietà notevoli, cioè S millepunctata, S tenuipunctata
e altre varietà quasi tipiche raggruppabili in una S subtipica, senza attribuzione di sigle sintetiche alle 3 entità
citate, tranne l’attribuzione specifica comune di S, mentre ho separato
un’ulteriore varietà notevole denominata “lellia” = F,
a sua volta suddivisa nelle 2 sub varietà “tondapex” e “acutapex”, queste senza sigle
distintive.
velai…………………….……...….W Galindo e Cossignani, 2020 (MMM n°108…)
visayae…………………..….…….V Mattavelli, 2021 (MMM n°113…). Esistono
almeno 2 varietà di V: tipica a punti ultra fini, compreso l’olotipo, e varietà pseudovisayae a
punti grossolani, che è stata poi siglata
Y.
Ecco inoltre l’elenco delle altre 9 presumibili “sottospecie” non ancora ben definite
come tali, o di diverso inquadramento
biologico da individuare, ma già morfologicamente definite, riservando la
stringa …X… alla composizione di presumibili effettivi ibridi sterili, o talora
forse klepton ibridogenici:
glabella? albida……………….B
Mattavelli 2005/2021 (v. Albinismo delle simil-glabella)
glabella? atalanta……………N Mattavelli 2021 (nel link Foto 12 B).
glabella? bordeaux………….U Conchology. Inc. (v. “ghepardiche”)
probabile varietà di C
sebastiani? galindoi?.........Z Mattavelli 2021 (Revisione di G Foto 4 e Albinismo_2 Fig. 16)
pseudoglabella? lellae……..E
Mattavelli 2018 (rif. ipotesi per M. visayae)
sebastiani? lellia…………..….F
Mattavelli 2006/2022 = varietà di S o improbabile klepton
glabella? neoglabella……...C Mattavelli 2021 (Revisione di G Foto 9 & 9B)
irrorata? pseudoirrorata….T
Mattavelli 2017 (Marginella
irrorata pseudoirrorata)
irrorata? tizianica…………...H
Mattavelli 2021 (Revisione di G Foto
18)
Nell’articolo 5 confondibili marginelle simil-glabella avevo adottato i simboli X, A, Y
e Z con significati diversi da quelli sopraesposti, Vi prego di dimenticare
quei significati, a favore degli attuali.
Nelle celle e nell’elenco finale delle specie
dell’articolo Revisione di M. glabella
compaiono dei morfi qui dimenticati, es. M.
glabella xantica, M. irrorata albinea. Probabilmente questi
morfi possono rientrare in varietà sottospecifiche già sopra elencate, occorre
verificarne la trascurabilità biologica.
Alcuni nomi da me introdotti in passato sono oggi inquadrabili in
specie/varietà meglio definite.
Es. i morfi “incredibilis”
del 2011 già poco oltre il 2012 venivano riqualificati M. pseudodesjardini;
altro es. “lumperia” oggi può rientrare in M. pseudoglabella. Avevo
già posto l’argomento “varietà lellia” nella pagina Universo lumperia, per poi
arrivare a definire M. pseudoglabella. In tale pagina sono presenti anche altre foto di “lellia” etc.
Marginella glabella antinea,
problematica, mutabilis oggi possono rientrare in M. (kl.?) visayae.
Altri nomi di varietà più uniche che rare sono
tuttora validi, ma biologicamente poco influenti, mentre sarebbe molto più
utile stabilire in quale modo possono esistere vere marginelle
klepton. Ad es., nell’articolo Altri klepton nel Marginella glabella Complex?
vedere i morfi “giselica
ed eurekia”.
Home, in 4 Sezioni: Conchiglie, Macchine, Aeromodellismo, Gorgonzola.
Tot. 53 pagine: Indice generale.
Indice solo della
Sezione Conchiglie (24 pagine).
Articoli amatoriali su argomenti riguardanti il complesso di marginelle ricollegabili alla specie Marginella
(Voluta) glabella Linneo,
1758/1767.
- Presentazione e prefazione della Sezione
Conchiglie
- Concetti di Specie/klepton. Aggiornamento 2016.
- Atlante delle specie/klepton attuali del Marginella glabella
Complex, 2022.
- Prima parte
dell’articolo La variabilità fatta specie, 2005, per la distinzione tra M. glabella e M. irrorata.
- Seconda pagina del sopracitato articolo, per gli ibridi morfologici M. glabella X irrorata: M. glabella
mutabilis, 2005, pagina di archivio da rivedere come M. visayae, 2021.
- Allegato alla Variabilità fatta specie, inerente M. glabella
antinea & problematica, 2005.
- Articolo Altri klepton nel Marginella glabella
Complex? con tabella
fotografica del Complex, 2021.
- Marginella pseudodesjardini
Le Béon, 2012 , con note su "pseudodesjardini giselica",
2013.
- Universo lumperia. Morfi ibridi e “specie” Marginella
(kl.?)
pseudoglabella, supposizione 2014.
- Note inerenti M. irrorata varietà pseudoirrorata
Mattavelli,
luglio 2017.
- Integrazione pagina Links al Complex, con
Marginella (kl.?) pseudoglabella, ipotesi 2017 .
- Articolo Nuovo sguardo ai mondi di M. glabella & pseudoglabella,
agosto 2017.
- Descrizione comparativa della nuova
“specie” Marginella (kl.?) pseudoglabella Mattavelli, 2018.
- Marginella (Kl.?) visayae Mattavelli, 2018 in Internet ipotizzata nuova
"specie" (kl. M. glabella X irrorata?), descritta
insieme a M. pseudoglabella lellae & M. irrorata pseudoirrorata,
pure ipotizzabili come “specie”, tuttavia ritenute solo “varietà” di
specie.
- 5 confondibili marginelle
simil-glabella, irrorata, velai, lamarcki e
visayae, relaise 2021.
- “Albinismo” delle simil
M. glabella
& irrorata, con ipotesi
di 2 nuove “specie albine” (M. albida & M.
tizianica Mattavelli 2021), descriziome solo in Internet, novembre 2021.
- ”Albinismo” in generale, anche per specie non del M. glabella
Complex, 2021.
- Revisione di M. glabella
Linneo, 1758, con mie nuove denominazioni di sue
varietà (es. M. glabella
glabella & M. glabella
neoglabella, 2021), solo in Internet, 2021.
- Marginella (kl.?) visayae Mattavelli, 2021 nuova
“specie sorella” di M. glabella & irrorata, a completamento della
descrizione cartacea, fatta sulla rivista MMM n° 113.
- Un salto nei retini conchigliari
“ghepardici”.
- Marginella sebastiani
nelle sue 5 varietà principali.
Articoli
della Sezione Conchiglie solo in Inglese:
- Atlas of the
Complex, ex Quick
reference guide (Update 2022).
- Marginella pseudosebastiani
Mattavelli 2001, statements versus M. sebastiani
(archivio 2006).
- Complex links, with Marginella (kl.?) pseudoglabella Mattavelli 2018 new sp.
. 2020 remake.